Farbwahrnehmung
Karl R. Gegenfurtner
Abteilung Allgemeine Psychologie
Justus-Liebig-Universität Gießen
Otto-Behaghel-Str. 10F
35394 Gießen
Telefon: +49 641 9926100
Fax: +49 641 9926119
e mail: gegenfurtner@uni-giessen.de
Farbe wird üblicherweise definiert als diejenige Empfindung, die es uns
ermöglicht, zwischen zwei strukturlosen Flächen gleicher Helligkeit zu unterscheiden.
Es lohnt sich, auf zwei Aspekte dieser Definition gleich zu Anfang näher
einzugehen. Zunächst ist wichtig, daß Farbe eine Empfindungsgröße ist. Es
ist nicht das Licht, das farbig ist ("The rays are not coloured" - Isaac Newton).
Das Licht wird zunächst im Auge in Nervenimpulse gewandelt. Erst durch die
Verarbeitung dieser Impulse in den nachgeschalteten Hirnstrukturen kommt
es zu der Empfindung, die wir “Farbe” nennen. Der zweite wichtige Aspekt
der obigen Definition betrifft die Funktion des menschlichen Farbensehens.
Obwohl die Frage nach der evolutionären Funktion des Farbensehens noch
heftig umstritten ist (Mollon & Jordan, 1988), läßt sich doch mit Sicherheit
sagen, daß diese nicht hauptsächlich in der Unterscheidung von Flächen
gleicher Helligkeit liegt. Zum einen tauchen solche rein spektralen Unterschiede
von Oberflächen in unserer Umgebung nur äußerst selten auf. Zum anderen
konnte in zahlreichen Untersuchungen in den letzten 20 Jahren gezeigt werden,
daß die Unterscheidung von Flächen gleicher Helligkeit (“Isoluminanz”) für
das menschliche visuelle System eine relativ schwierige Aufgabe darstellt
(Shapley, 1990).



Abb. 1: Photographie einer Szene mit Blumen.
Links das Original, in der Mitte eine Schwarz-weiß-Version, rechts eine
isoluminante Version, bei denen allen Bildpunkten dieselbe Helligkeit zugeordnet
wurde.
Im folgenden wird zuerst auf die physikalischen
Eigenschaften der Reize eingegangen, die im menschlichen visuellen System
Farbempfindungen auslösen. Anschließend werden die wichtigsten Schritte
der Verarbeitung in der Netzhaut durch die drei Typen von Photorezeptoren
und in den anschließenden Gegenfarbkanälen besprochen. Schließlich wird
auf die Verarbeitung der Farbinformation im visuellen Kortex eingegangen.
Während die ersten Stufen der Farbverarbeitung in der Netzhaut und den retinalen
Ganglienzellen wohl besser als jeder andere Aspekt der visuellen Wahrnehmung
erforscht sind, ist über höhere Verarbeitungsmechanismen, die semantische
oder emotionale Aspekte der Farbe betreffen, bisher nur wenig bekannt. 1. Retinale Verarbeitung
1.1. Licht und Farbe
Die physikalische Grundlage
für die Farbwahrnehmung ist elektromagnetische Strahlung mit einer Wellenlänge
in einem eng umgrenzten Bereich von ca. 400 bis 700 Nanometer. Strahlung
in diesem Wellenlängenbereich ist nur deshalb sichtbar, weil wir Rezeptoren
in der Netzhaut des Auges besitzen, deren Pigmente durch Licht chemisch
verändert werden können. Sie unterscheidet sich ansonsten in keiner Weise
von anderen Strahlungen, wie z.B. Radiowellen, Infrarot-, Ultraviolett-,
Röntgen- oder Gammastrahlen. Ein Großteil der Sonnenstrahlung, die die
Erdoberfläche erreicht, liegt im sichtbaren Bereich. Daher ist anzunehmen,
daß sich unser Farbsehsystem im Laufe der Entwicklung optimal an die Gegebenheiten
unserer Umwelt angepaßt hat.
Das Spektrum des in das Auge
fallenden Lichts hängt von zwei Faktoren ab. Von der Spektralverteilung
der Beleuchtungsquelle, und von der Reflektanz der Objekte, über die das
Licht ins Auge reflektiert wird. Nur ein Teil des auf ein Objekt fallenden
Lichts wird reflektiert. Der andere Teil der Strahlung wird absorbiert und
zumeist in Wärmeenergie umgewandelt. Dunklere Objekte absorbieren mehr Licht
und erwärmen sich daher stärker. Das ins Auge gelangende Licht ist das Produkt
aus Beleuchtung und Reflektanz, so daß diese beiden Faktoren anschließend
nicht mehr getrennt werden können. Ein rötlicher Farbeindruck kann danach
gleichermaßen durch rötliche Beleuchtung eines weißen Stück Papiers, oder
durch weiße Beleuchtung eines rötlichen Stück Papiers zustandekommen. Obwohl
mathematisch beides zur gleichen Wellenlängenverteilung führt, kann unser
visuelles System zumeist doch Rückschlüsse auf die Reflektanz der Objekte
ziehen. Diese Leistung, die durch die Ambiguität von Beleuchtung und Reflektanz
notwendig wird, wird “Farbkonstanz” genannt. Der Vorteil der Farbkonstanz
liegt darin, daß die Reflektanz eine invariante Eigenschaft der Objekte
ist, und sich daher zur Objekterkennung sehr gut eignet.
Farbe ist nicht gleich
Wellenlänge. Während monochromatisches Licht zwar unter neutralen Beobachtungsbedingungen
immer die gleiche Farbempfindung auslöst, ist die umgekehrte Zuordnung nicht
möglich. Natürlich auftretende Objekte reflektieren immer Licht über einen
weiten Bereich von Wellenlängen. Der relative Anteil in den verschiedenen
Wellenlängenbereichen bestimmt letztendlich, welche Farbe gesehen wird. Des
weiteren kann das Umfeld auch massive Einflüsse auf die Farbwahrnehmung haben.
1.2. Verarbeitung in den Photorezeptoren
Im Auge wird das Licht von
Photorezeptoren absorbiert. Es gibt zwei Klassen von Photorezeptoren: Stäbchen
und Zapfen. Stäbchen sind sehr lichtempfindlich und ermöglichen das Sehen
bei Dunkelheit und in der Dämmerung. Da alle Stäbchen dieselbe spektrale
Absorption aufweisen, können sie nicht zwischen Wellenlängen- und Intensitätsunterschieden
diskriminieren. Farbunterscheidungen sind daher beim Stäbchensehen nicht
möglich. Unter Tageslichtbedingungen sind die Stäbchen vollständig gesättigt
und unfähig Information zu verarbeiten. Dann sind die weniger lichtempfindlichen
Zapfen aktiv.
Eine ganz besondere Form der
genetisch bedingten, totalen Farbenblindheit ist die Stäbchenmonochromasie.
Dabei sind nur Stäbchenphotorezeptoren in der Netzhaut vorhanden, was zu
einer ganzen Reihe von Symptomen wie z. B. Photophobie, einer extrem schlechte
Sehschärfe und massiven Fixationsproblemen führt. Diese Form der Farbenblindheit
wurde von Oliver Sacks (1997) in dem Roman “Insel der Farbenblinden”
beschrieben. Eine beeindruckende, subjektive Darstellung aus der Sicht
eines Stäbchenmonochromaten findet sich auch in Nordby (1990).
Von den Zapfenphotorezeptoren
gibt es drei verschiedene Arten. Nach dem Spektralbereich ihrer höchsten
Empfindlichkeit werden sie lang-, mittel-, und kurzwellenlängen-sensitiv
genannt, oft auch nur kurz Rot-, Grün- und Blauzapfen. In Abbildung 2 sind
die Absorptionsspektren dieser drei Zapfenarten dargestellt. Man erkennt,
daß alle drei Typen über einen großen Wellenlängenbereich hinweg Licht absorbieren.
Die Kurzbezeichnungen sind also irreführend, vor allem wenn man berücksichtigt,
daß das Maximum für die Rotzapfen in dem Wellenlängenbereich liegt, den wir
als gelb wahrnehmen. Das Maximum der Blauzapfen liegt in einem Bereich, der
eher violett aussieht als blau. Da sich diese Bezeichnungen aber eingebürgert
haben, werden wir sie hier der Einfachheit halber auch beibehalten. Es muß
aber nachdrücklich betont werden, daß andere mit R, G, B bezeichnete Farbsysteme,
wie z.B. das C.I.E. RGB System, oder auch die Rot-, Grün- und Blauphosphore
von Farbbildschirmen nichts oder nur sehr wenig mit den Absorptionseigenschaften
der Zapfen gemeinsam haben.
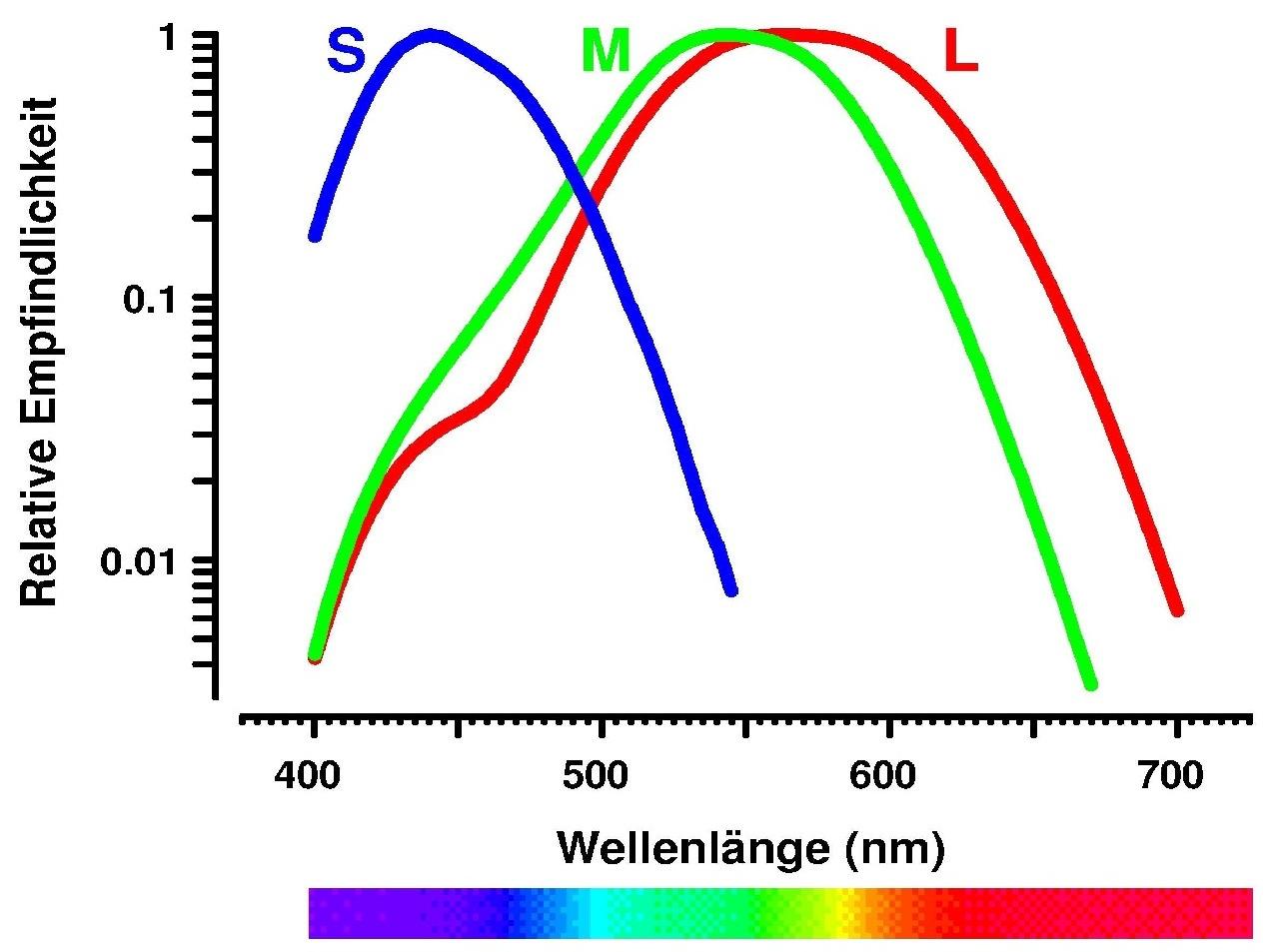
Abb. 2: Spektrale Absorptionskurven der menschlichen
Rot-, Grün- und Blauzapfen.
Aus Abbildung 2 ist auch ersichtlich, daß die Absorptionsspektren
für die Rot- und Grünzapfen sehr ähnlich sind. Die Absorptionsgipfel sind
nur um ca. 30 Nanometer verschoben. Dies hat evolutionäre Gründe: diese zwei
Zapfentypen sind erst vor entwicklungsgeschichtlich relativ kurzer Zeit
aus einem gemeinsamen Urzapfen entstanden. Die Absorption der Zapfen hängt
von ihrem Sehfarbstoff ab, dessen Proteine genetisch bestimmt sind. Jeremy
Nathans und seinen Mitarbeitern ist es gelungen, die Gene zu identifizieren,
die die Ausbildung dieser Proteine kodieren (Nathans, 1992). Dabei hat sich
herausgestellt, daß sich die Aminosäuresequenzen für das Rot- und das Grünpigment
nur an wenigen Stellen unterscheiden (< 2%).
Aus der Tatsache, daß die Zapfen das Farbensehen
ermöglichen, wird oftmals der falsche Schluß gezogen, daß die Stäbchen für
das Helligkeits- oder Schwarz-weiß-Sehen zuständig sind. Das stimmt nicht!
Unter Tageslichtbedingungen liefern die Stäbchen keinerlei brauchbare Signale.
Sie sind nur beim Dämmerungs- und Nachtsehen, z.B. bei Mondlicht, aktiv.
| Geschlecht |
Anzahl |
Rot-Zapfen (Protan) |
Grün-Zapfen (Deutan) |
||
| |
|
Anomalie |
Anopia |
Anomalie |
Anopia |
| männlich |
45,989 |
1.08 |
1.01 |
4.63 |
1.27 |
| weiblich |
30,711 |
0.03 |
0.02 |
0.36 |
0.01 |
Tabelle 1: Relative Häufigkeiten (in %) von
genetisch bedingten Rot-grün Farbsehsörungen. Rot-grün- Blindheit (Anopia)
taucht bei ca. 2.3% der männlichen Bevölkerung Europas auf. Rot-grün-Störungen
(Anomalie) treten bei weiteren ca. 5.7% auf (nach Sharpe et al., 1999, Tabelle
1.4).
1.3. Die Netzhaut
Die Zapfen sind auf der Netzhaut zu
einem unregelmäßigen Mosaik angeordnet. Die Dichte ist in der Fovea am höchsten
und nimmt zur Peripherie hin ab. In der Foveola (den zentralen 30') befinden
sich nur Rot- und Grünzapfen auf. Blauzapfen gibt es nur in der peripheren
Retina, aber auch dort treten sie mit einer geringeren Dichte auf. Sie
machen insgesamt nur 9% aller Zapfen aus. Da die Sehschärfe von der Dichte
der Zapfen abhängt, ist die Auflösung für Muster, die gezielt Blauzapfen
anregen, relativ gering. Diese geringere Auflösung wiederum scheint aber
perfekt an die Optik des Auges angepaßt zu sein. Da Licht unterschiedlicher
Wellenlänge wegen der unterschiedlich starken Brechung nicht gleichzeitig
auf der Netzhaut fokussiert werden kann, entsteht vor allem bei kurzwelligem
Licht eine retinale Unschärfe, die dem größeren Abstand zwischen den Blauzapfen
entspricht. Das Zapfenmosaik ist auch in anderer Hinsicht sehr gut der Optik
des Auges angepaßt. Dort wo die Zapfendichte geringer ist als aufgrund des
optischen Signals notwendig (in der Peripherie), sorgt die unregelmäßige
Anordnung der Zapfen dafür, daß keine Wahrnehmungstäuschungen (durch “Aliasing”)
entstehen.

Abb. 3: Simulation des Zapfenmosaiks der Fovea. Die Positionen der Zapfen entstammen anatomischen Messungen. Die Einfärbung wurde nach einem Zufallsschema durchgeführt unter der Annahme, daß Rotzapfen ca. doppelt so häufig sind wie Grünzapfen. [Die Abbildung wurde freundlicherweise von Herbert Jägle und Ted Sharpe von der Augenklinik Tübingen zur Verfügung gestellt.
Die Frage nach der relativen
Anzahl von Rot- und Grünzapfen hat die Farbforscher in den letzten zehn
Jahren intensiv beschäftigt, wobei die Ergebnisse verschiedener Methoden
zu höchst unterschiedlichen Ergebnissen geführt haben. Vor kurzem gelang
es jedoch Roorda & Williams (1999), das Photorezeptormosaik der menschlichen
Netzhaut direkt abzubilden (siehe Abb. 3). Es zeigten sich große Unterschiede
zwischen verschiedenen Probanden, und auch letztendlich, daß das Verhältnis
von Rot- und Grünzapfen keinerlei Einfluß auf subjektive Farbeindrücke besitzt.
Es wird oftmals behauptet, daß im peripheren Gesichtsfeld keine Farbwahrnehmung möglich ist. Zu Demonstrationszwecken wird dann ein buntes Objekt von der Fovea weg bewegt, bis die Farbe des Objekts nicht mehr erkannt wird. Dies zeigt jedoch nur, daß die räumliche Auflösung des Farbsehsystems geringer ist als die des Helligkeitssystems. Wenn die Objekte nur ausreichend groß sind, dann kann auch deren Farbe bei peripherer Darbietung erkannt werden.
1.4. Gegenfarben
Die von den Zapfen übermittelten Signale werden
noch in der Netzhaut weiterverarbeitet, um die Weiterleitung der Signale
in den visuellen Kortex möglichst optimal zu gestalten. Diese Verarbeitungsstufe
läßt sich am besten als Verminderung der Redundanz der Aktivitäten benachbarter
Zapfen charakterisieren. Die Zapfensignale weisen sowohl eine räumliche
als auch eine farbliche Redundanz auf.
Räumliche Redundanz bezeichnet den Sachverhalt,
daß benachbarte Bildpunkte meistens eine ähnliche Intensität aufweisen.
Aus der Intensität an einem Bildpunkt läßt sich relativ genau die Intensität
der benachbarten Bildpunkte vorhersagen. Werden hingegen die Differenzen
zwischen benachbarten Bildpunkten betrachtet, so verschwindet diese Korrelation.
Diese Art der vom visuellen System durchgeführten Differenzenbildung wird
oftmals auch als “laterale Hemmung” bezeichnet. Implementiert ist sie durch
die konzentrischen rezeptiven Felder der Ganglienzellen, in denen Zentrum
und Umfeld antagonistisch organisiert sind.
Eine weitere Form der Redundanz in den Zapfen
ergibt sich daraus, daß die Absorptionsspektren der Rot- und Grünzapfen
sehr ähnlich sind. Daher ist die Aktivität der beiden Zapfentypen hoch korreliert.
Um diese Signale zu dekorrelieren und damit zu optimieren, wird die Aktivität
in sogenannten Gegenfarbkanälen, oftmals auch als “kardinale Farbrichtungen
bezeichnet, weitergeleitet. Im Helligkeitskanal wird die Summe der Signale
aus Rot- und Grünzapfen (R+G) gebildet, im Rot-Grün-Kanal die Differenz
der beiden (R-G). Im Blau-Gelb-Kanal schließlich wird die Differenz aus
dem Signal der Blauzapfen und der Summe der Rot- und Grünzapfen gebildet
(B‑(R+G)). Diese Verrechnung der Farbsignale erfolgt in einem Netzwerk aus
Horizontal-, Bipolar- und Ganglienzellen. Während über die Ergebnisse dieser
Berechnung relative Klarheit herrscht, ist die genaue Implementierung derzeit
Gegenstand heftiger Diskussion. Es scheint so zu sein, daß die drei Gegenfarbkanäle
nicht nur funktionell, sondern auch anatomisch unterschiedlich sind (siehe
Gegenfurtner & Sharpe, 1999).



Abb. 4: Verarbeitung in den Gegenfarbkanälen.
Links das Original, in der Mitte das Bild, wie es vom Rot-Grün-Kanal gesehen
wird. Rechts das Bild, wie es vom Blau- Gelb-Kanal gesehen wird.
Eine wichtige Implikation der Gegenfarbkanäle
ergibt sich aus der starken Überlappung der Absorptionsspektren von Rot-
und Grünzapfen. Es ist sehr schwierig, den Rot-Grün- Kanal so zu
aktivieren, daß die Differenz von Rot und Grün sich ändert, aber die Summe
(die Helligkeit) konstant bleibt. Die größtmögliche Modulation der Zapfensignale
bei einer solchen "isoluminanten" Reizung bleibt daher weit hinter der
100%-igen Modulation zurück, die durch Helligkeitskontraste bewerkstelligt
werden kann. Auf Bildschirmen beträgt diese maximale isoluminante Modulation
bei mittleren Intensitäten nur ca. 10-15%! An dieser Stelle ist es auch
wieder wichtig, auf den Unterschied zwischen Zapfen und Bildschirmphosphoren
hinzuweisen. Letztere können nämlich auch bei Isoluminanz zu 100% moduliert
werden. Für das Studium des visuellen Systems ist das allerdings irrelevant.
Die Einteilung der Signalverarbeitung
in zwei Zonen, der anfänglichen Verarbeitung in drei verschiedenen Zapfentypen,
gefolgt von drei Gegenfarbkanälen, vereint die Theorien der Farbwahrnehmung
von Helmholtz und Hering. Allerdings hatten beide dieser eminenten Forscher
nicht ganz recht, was die Details betrifft. So nahm Helmholtz an, daß die
Absorptionsspektren der Zapfen sich nur geringfügig überlappen, was für die
Rot- und Grünzapfen gerade nicht zutrifft. Hering nahm an, daß die Gegenfarben
den sogenannten Urfarben entsprechen, also denjenigen Farben, die von uns
als reines Rot, Grün, Blau, oder Gelb wahrgenommen werden. Auch dies ist
nicht richtig. Während das Rot der “kardinalen Farbrichtungen” der Ganglienzellen
in etwa einem Urrot entspricht, sieht die Gegenfarbe dazu blau-grün aus.
Der Blau-Gelb-Kanal ist ebenfalls verschoben: dem reinem Gelb entspricht ein
grünlicher Gelbton und dem Blau ein Violett. Abbildung 4 zeigt wie die Gegenfarbkanäle
aktiviert werden.
Für die folgende Diskussion
der Wahrnehmungsleistungen des Farb- und Helligkeitssystems ist es allerdings
sehr wichtig, solche peripheren Faktoren von zentralen Unterschieden in der
Verarbeitung zu trennen. Wir werden zunächst auf das Zusammenspiel von Farbe
mit anderen visuellen Reizattributen eingehen, dann auf die Verarbeitung
des Farbsignals an sich. Als Abschluß folgen einige Bemerkungen zur Farbkonstanz.
2. Kortikale Farbmechanismen
Ein allgemeines Prinzip kortikaler
Informationsverarbeitung ist eine zunehmende Spezialisierung der Neurone.
Dies trifft auch für die kortikale Verarbeitung der Farbe zu. In der Netzhaut
gibt es genau drei Klassen von Neuronen, die den kardinalen Farbrichtungen
entsprechen (siehe Abb. 4). Die Antworten dieser Neurone sind linear. Daher
läßt sich die Antwort auf beliebige Reize exakt vorhersagen durch den bevorzugten
Farbreiz eines Neurons. Im primären visuellen Kortex (V1) gilt dieses Prinzip
der Linearität auch, aber hier finden sich Präferenzen für beliebige Farben.
Die Einschränkung auf die drei kardinalen Richtungen entfällt. Interessant
ist, daß bisher kein neuronales Substrat für die sogenannten “Urfarben”
nachgewiesen werden konnte. Die Vielfalt an Farbpräferenzen, die in V1 und
V2 beobachtet werden kann, entspricht unserer Fähigkeit eine große Anzahl
an Farben unterscheiden zu können.
Im sekundären visuellen Kortex
(V2) treten dann auch Neurone auf, deren Reizantwort im spektralen Bereich
nichtlinear ist. Sie antworten nur auf einen bestimmten Farbton, da ihre
chromatische Bandbreite sehr eng ist. Zudem ist es in den höheren extrastriären
Arealen (V2, V3, V4) auch so, daß Neurone zunehmend seltener auf reine (isoluminante)
Farbunterschiede antworten. Da solche isoluminanten Unterschiede, wie schon
eingangs erwähnt, nicht sehr oft in unserer Umwelt vorkommen, wäre eine
Spezialisierung auf die Verarbeitung dieser Reize auch nicht sehr sinnvoll.
2.1. Das Farbzentrum im Gehirn?
Im Zentrum derzeitiger Forschung
auf dem Gebiet der Farbwahrnehmung steht wohl die Frage, ob es im Gehirn
eine bestimmte neuronale Strukur gibt, die vornehmlich für die Verarbeitung
von Farbinformation zuständig ist. Eine solche “parallele” Verarbeitung
wurde in der Vergangenheit auf Grund psychophysischer, physiologischer,
und klinischer Befunde postuliert (Livingstone & Hubel, 1988; Zeki,
1990).
So ist z. B. das Erkennen von
Formen und Strukturen bei ausschließlich durch Farbe definierten Reizen
sicherlich anders als bei Helligkeitsmustern (Livingstone & Hubel, 1988).
Dies läßt sich schon aus den oben erwähnten peripheren Faktoren folgern.
Der höchstmögliche Kontrast im Rot-Grün-Kanal ist wegen der großen Ähnlichkeit
von Rot- und Grünzapfen sehr stark eingeschränkt. Wegen optischer Limitationen
(chromatische Aberration) können hohe Ortsfrequenzen vom Farbsystem nicht
übertragen werden. Letzteres wird auch bei der Übertragung von Farbfernsehbildern
ausgenutzt, wo die Farbinformation mit einer sehr viel geringeren Bandbreite
gesendet wird. Viele der Demonstrationen, die eine getrennte Verarbeitung
von Farbe und anderen visuellen Reizattributen nahezulegen scheinen, beruhen
auf derartigen peripheren, retinalen, Unterschieden. Die physiologische
und anatomische Trennung in unterschiedliche Verarbeitungskanäle scheint
dabei weitaus geringfügiger zu sein als ursprünglich angenommen (siehe Lennie,
1998).
Studien mit bildgebenden Verfahren
zeigen oft eine erhöhte, farbspezifische Aktivierung in einem Bereich des
menschlichen ventralen Okzipitalkortex, der oftmals “V4” genannt wird (Zeki,
1990; Hadjikani et al., 1998; Engel, Zhang & Wandell, 1997). Die Aktivierung
scheint sich allerdings qualitativ nicht von der im primären visuellen
Kortex zu unterscheiden. Es kann also nicht geschlossen werden, daß in
dem entsprechenden Areal nur Farbinformation verarbeitet
wird, oder daß andere Areale keine wichtigen Beiträge zur Verarbeitung
von Farbinformation liefern. Interessanterweise handelt es sich dabei jedoch
um denselben Bereich, der auch oftmals bei Patienten betroffen ist, die
stark erhöhte Schwellen für Farbunterscheidung aufweisen (“zerebrale Achromasie”).
Viele dieser Patienten weisen jedoch noch weitere Wahrnehmungsstörungen
auf, z.B Störungen der Gesichtserkennung oder eine allgemein verschlechterte
Kontrastempfindlichkeit. “Reine” Störungen der Farbwahrnehmung sind selten
(Zihl & von Cramon, 1986).
2.2. Farbkonstanz
Ein Thema, das Wahrnehmungsforscher
und Informatiker in jüngster Zeit gleichermaßen stark interessiert hat,
ist die Farbkonstanz. Die Effekte von Beleuchtung und Reflektanz lassen
sich nicht trennen, da das auf die Rezeptoren fallende Licht das Produkt
beider Faktoren ist. Trotzdem scheint es uns möglich zu sein, Objekte unter
sich wechselnden Beleuchtungsbedingungen immer mit der gleichen Farbe wahrzunehmen.
Abbildung 5 zeigt eine Waldlandschaft unter drei verschiedenen Beleuchtungen,
wie sie z.B. morgens, mittags und abends aussehen könnte. Wenn wir durch
die Wiesen wandern, erscheint uns das Gras meist im selben Grünton, obwohl
der Farbeindruck auf den Bildern deutlich unterschiedlich ist.



Abb. 5: Darstellung einer natürlichen Szenen,
wie sie unter drei verschiedenen Beleuchtungsbedingungen aussieht. Die Veränderung
von morgens (links), mittags (mitte) und abends (rechts) erfolgt hauptsächlich
entlang der blau-gelben Gegenfarbachse.
Eine Unmenge an Algorithmen
wurde vorgeschlagen, um diese Farbkonstanz zu erklären. Mittlerweilen zeichnet
sich ab, daß das menschliche Sehsystem eine ganze Reihe von Hinweisreizen
benutzt um Farbkonstanz zu erzielen (Kraft & Brainard, 1999). Am wichtigsten
ist dabei die Tatsache, daß die Mittelwertsfarbe in einer Szene sehr stark
von der Beleuchtung abhängt, und daß Beleuchtungsänderungen meistens graduell
vonstatten gehen, während Reflektanzänderungen oft abrupt sind.
Dieser Unterschied in den räumlichen
Eigenschaften von Beleuchtung und Reflektanz kann am besten genutzt werden,
wenn größere Flächen für eine Normalisierung benutzt werden. Dies erfordert
daher rezeptive Felder mit entsprechender Größe. Neurone mit diesen Eigenschaften
wurden in V4 gefunden, und sie scheinen auch relativ komplexe Interaktionen
von Zentrums- und Umfeldfarbe aufzuweisen. Wie daraus aber dann die Reflektanz
von Objekten bestimmt wird, ist noch weitgehend unklar. Neurone, die auf
Reflektanz antworten, wurden bislang nur selten und vereinzelt gefunden
(Zeki, 1980).
Störungen der Farbkonstanz
wurden bislang in erster Linie an Probanden untersucht, die bereits ander
massive Farbsehstörungen aufweisen. In den Arbeiten von Rüttiger et al (1999)
und Clarke et al. (1999) sind aber Patienten beschrieben, deren Farbkonstanzleistungen
gestört sind, obwohl sie keinerlei andere erkennbaren Störungen in ihrer
Farbwahrnehmung aufwiesen. Über das neuronale Substrat der Farbkonstanzleistung
läßt sich noch keine Schlußfolgerung ziehen.
2.3. Kognition, Emotion und Farbe
In der weiteren kognitiven
Verarbeitung der Farbinformation werden verschiedene Farbnuancen zu Kategorien
zusammengefaßt. Von diesen Kategorien gibt es ca. 7 bis 11, die sogar über
größere Kulturunterschiede hinweg relativ konstante Bezeichnungen aufweisen
(siehe Hardin & Maffi, 1997). Ein neuronales Substrat für derartige Kategorien
wurde bislang noch nicht entdeckt. Zumindest in den frühen visuellen Verarbeitungsstufen,
bis hin zum extrastriären Areal V4, findet sich keine bevorzugte Repräsentation
dieser Kategorien. Die Zuweisung von Farbnamen zu Objekten scheint daher
auf einer sehr hohen Verarbeitungsebene abzulaufen, während einfache Farbunterscheidungen
schon durch Schaltkreise im primären visuellen Kortex erklärt werden können.
Dies wird auch durch Ergebnisse aus der Entwicklung belegt. Kinder können
Farben schon im Alter von ca. 4 Monaten unterscheiden, während die richtige
Benennung erst sehr viele später, im Alter von 2-4 Jahren erlernt wird
(Bornstein, 1985).
Noch wichtiger als kognitive
erscheinen die emotionalen Aspekte, die oftmals mit der Wahrnehmung von
Farben einhergehen. Eine enge Vernetzung der kortikalen Farbsehmechanismen
mit limbischen Strukturen kann daher angenommen werden. Der Großteil dieser
emotionalen Reaktionen ist sicherlich erlernt und hängt von kulturellen Faktoren
ab. Es scheint aber auch einige universelle Phänomäne zu gebe. So geht die
Farbe “rot” generell einher mit gesteigerten emotionen Reaktionen, und ist
in nahezu allen Kulturen der erste, wichtigste Farbname.
Zusammenfassung
Farbe wird wahrgenommen wenn
Licht im Auge von Photorezeptoren absobiert und in Nervenimpulse umgewandelt
wird, die dann im Gehirn zu Empfindungen interpretiert werden. In der Netzhaut
unterliegen drei verschiedenen Typen von Zapfenphotorezeptoren, die jeweils
über weite Bereiche des Spektrums empfindlich sind, der Farbwahrnehmung.
Die von den Zapfen vermittelten Signale werden noch in der Netzhaut in den
retinalen Ganglienzellen zu den effizienteren Gegenfarbsignalen umkodiert,
von denen es wiederum drei Klassen gibt. Im visuellen Kortex entsteht dann
aus diesen Signalen eine Vielfalt von Mechanismen, die ganz spezifisch für
bestimmte Farbkombinationen empfindlich sind. Auf diesen Mechanismen basieren
Fähigkeiten wie z.B. Farbunterscheidung oder Farbkonstanz. Auf einer höheren,
kognitiven Ebene werden aus den Farben dann eine kleine Anzahl von Kategorien
(ca. 7-11) gebildet, die in nahezu allen Kulturen und Sprachen gleich sind.
Die wichtigsten Ursachen für Störungen der Farbwahrnehmung sind der genetisch
bedingte Ausfall eines Zapfentypen, und erworbene Farbenblindheiten, die
auf kortikalen Läsionen beruhen.
Literaturverzeichnis
Bornstein,
M.H. (1985) On the development of color naming in young children: data
and theory. Brain & Language, 26, 72-93.Clarke, S., Walsh, V., Schoppig, A., Assal, G. & Cowey, A. (1998) Colour constancy impairments in patients with lesions of the prestriate cortex. Experimental Brain Research, 123, 154-158.
Engel, S.A., Zhang, X., & Wandell, B.A. (1997) Color tuning in human visual cortex measured using functional magnetic resonance imaging. Nature, 388, 68–71.
Gegenfurtner, K.R. & Rieger, J. (2000) Sensory and cognitive contributions of color to teh recognition of natural scenes. Current Biology, 10, 805-808.
Gegenfurtner, K.R. & Sharpe, L.T. (1999) Color vision: from genes to perception. Cambridge University Press, New York.
Hadjikani, N., Liu, A.K., Dale, A.M., Cavanagh, P. & Tootell, R.B.H. (1998) Retinotopy and color sensitivity in human visual cortical area V8. Nature Neuroscience, 1, 235-241.
Hardin, C.L. & Maffi, L. (1997) Color categories in thought and language. Cambridge University Press, New York.
Kraft, J.M. & Brainard, D.H. (1999) Mechanisms of color constancy under nearly natural viewing. Proceedings of the National Academy of Sciences of the United States of America, 96, 307-312.
Nordby, K. (1990) Vision in a complete achromat: a personal account. In Night Vision: Basic, Clinical and Applied Aspects (eds. Hess, R.F., Sharpe, L.T., & Nordby, K.), Chapter 7, pp. 290–315. Cambridge University Press, Cambridge.
Lennie, P. (1998) Single units and visual cortical organization. Perception, 27, 889–935.
Livingstone M.S. & Hubel D. (1988) Segregation of form, color, movement, and depth: anatomy, physiology, and perception. [Review]. Science, 240, 740-749.
Mollon, J.D. & Jordan, G. (1988) Eine evolutionäre Interpretation des menschlichen Farbensehens. Die Farbe, 35/36, 139–170.
Nathans, J. (1992) Die Gene für das Farbensehen. In W. Singer (Hrgb.) Gehirn und Kognition. Spektrum Akademischer Verlag, Heidelberg.
Roorda, A. & Williams, D.R. (1999) The arrangement of the three cone classes in the living human eye. Nature, 397, 520–522.
Rüttiger, L., Braun, D.I., Gegenfurtner, K.R., Petersen, D., Schönle, P. & Sharpe, L.T. (1999) Selective colour constancy deficits after circumscribed unlilateral brain lesions. Journal of Neuroscience, 19, 3094-3106.
Sacks, O. (1997) The Island of the Colorblind. Knopf, New York.
Shapley, R. (1990) Visual sensitivity and parallel retinocortical channels. Annual Review of Psychology, 41, 635–658.
Sharpe, L.T., Stockman, A., Jägle, H. & Nathans, J. (1999) Opsin genes, cone photopigments, color vision, and color blindness. In: Gegenfurtner, K.R. & Sharpe, L.T. (Hrg) Color vision: from genes to perception, pp 3-51. New York, Cambridge University Press.
Zeki S. (1980) The representation of colours in the cerebral cortex. Nature, 284, 412-418.
Zeki, S. (1990) A century of cerebral achromatopsia. Brain, 113, 1721–1777.
Zihl, J. & von Cramon, D. (1986) Zerebrale Sehstörungen. Stuttgart, Kohlhammer.